2.3 Brain activity measurement during speech
Brain function during speech production has been investigated by function
MRI. The view of brain regions involved in speech production has been
updated from the classical idea centered at Brocaユs area: currently
more attention is paid to the left anterior insula because this region
is selectively involved in the disorder of speech motor programming.
However, functions of the insula are not fully understood, partly because
this region is located beneath the central sulcus. Following clinical
studies, many researchers conducted functional imaging experiments to
show evidence of involvement of the insula in speech production.
Our results as well as those of others were not always consistent,
however; the insula was often silent in repetitive tasks and tended
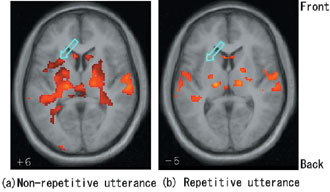
to be more active in non-repetitive tasks. Figure 3 shows an example
of the data that were obtained while the task (three-syllable word)
was renewed at each repetition of the utterance. It appears that in
repetitive tasks, the processing from sound patterns to motor programs
is done only once at the first utterance, while in non-repetitive tasks,
it is done at each utterance. In other words, phonological representation
of the syllable sequence is decoded into motor patterns when a new task
is presented [3]. Our tentative conclusion is that the decoding of a
syllable sequence may be carried out in the left anterior insula.
References
[1] Shimada, Y., Fujimoto, I., Takemoto, H., Takano, S., Honda, K.,
& Takeo, I. (2002). 4D-MRI using synchronized sampling method (SSM).
Japanese Journal of Radiological Technology 58 (12), 1592-1598.
[2] Honda, K., Takemoto, H., Kitamura, T., Fujita, S., & Takano, S.
(2004). Exploring human speech production mechanisms by MRI. IEEE Trans.,
Inf., & Syst., E87-D, 1050-1058.
[3] Nota, Y., & Honda, K. (2003). Possible role of the anterior insula
in articulation. Proc. 6th Int. Seminar on Speech Production, 191-194.